Biosynthesis colesterol a'i fiocemeg - Diabetes
Heb amheuaeth, colesterol yw'r lipid mwyaf adnabyddus i'r cyhoedd; mae'n enwog oherwydd y gydberthynas uchel rhwng colesterol gwaed uchel ac amlder clefydau cardiofasgwlaidd dynol. Rhoddwyd llai o sylw i rôl hanfodol colesterol fel cydran o bilenni celloedd ac fel rhagflaenydd hormonau steroid ac asidau bustl. Mae colesterol yn angenrheidiol ar gyfer llawer o anifeiliaid, gan gynnwys bodau dynol, ond mae ei bresenoldeb mewn bwyd mamalaidd yn ddewisol - gall celloedd y corff eu hunain ei syntheseiddio o ragflaenwyr syml.
Mae strwythur y cyfansoddyn 27-carbon hwn yn awgrymu llwybr cymhleth ar gyfer ei biosynthesis, ond darperir ei holl atomau carbon gan ragflaenydd sengl - asetad. Blociau isoprene - y canolradd pwysicaf o asetad i golesterol, maent yn rhagflaenwyr llawer o lipidau naturiol, ac mae'r mecanweithiau ar gyfer polymeru blociau isoprene yn debyg ym mhob llwybr metabolaidd.
Dechreuwn trwy archwilio'r prif gamau yn llwybr biosynthesis colesterol o asetad, yna trafod cludo colesterol trwy'r llif gwaed, ei amsugno gan gelloedd, rheoleiddio arferol synthesis colesterol, a rheoleiddio mewn achosion o amsugno neu gludiant â nam. Yna edrychwn ar sylweddau eraill sy'n dod o golesterol, fel asidau bustl a hormonau steroid. Yn olaf, bydd disgrifiad o'r llwybrau biosynthetig ar gyfer ffurfio llawer o gyfansoddion - deilliadau o flociau isoprene, lle mae camau cynnar cyffredin gyda synthesis colesterol, yn dangos amlochredd rhyfeddol anwedd isoprenoid mewn biosynthesis.
Cynhyrchir colesterol o asetyl-CoA mewn pedwar cam
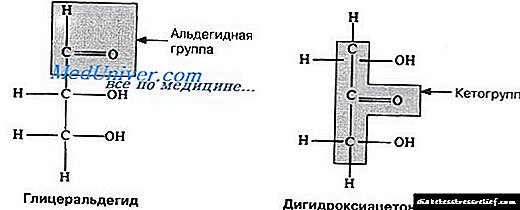
Mae colesterol, fel asidau brasterog cadwyn hir, wedi'i wneud o asetyl-CoA, ond mae'r patrwm cydosod yn hollol wahanol. Yn yr arbrofion cyntaf, ychwanegwyd asetad wedi'i labelu â 14 C naill ai yn yr atom carbon methyl neu garboxyl at fwyd anifeiliaid. Yn seiliedig ar ddosbarthiad y label mewn colesterol wedi'i ynysu oddi wrth ddau grŵp o anifeiliaid (Ffig. 21-32), disgrifiwyd camau ensymatig biosynthesis colesterol.
Ffig. 21-32. Ffynhonnell atomau carbon colesterol. Wedi'i nodi yn ystod arbrofion gan ddefnyddio asetad ymbelydrol wedi'i labelu â charbon methyl (du) neu garbonyl carbon (coch). Yn y strwythur cyddwys, dynodir modrwyau gan lythrennau A i D.

Mae'r synthesis yn digwydd mewn pedwar cam, fel y dangosir yn Ffig. 21-33: (1) cyddwysiad tri gweddillion asetad i ffurfio canolradd chwe charbon o mevalonate, (2) trosi mevalonate yn flociau isoprene actifedig, (3) polymerization chwe uned isoprene pum carbon i ffurfio squalene llinellol 30-carbon, (4) cylchredeg squalene i ffurfio pedair cylch o'r niwclews steroid, ac yna cyfres o newidiadau (ocsideiddio, tynnu neu fudo grwpiau methyl) wrth ffurfio colesterol.
Ffig. 21-33. Llun cyffredinol o biosynthesis colesterol. Trafodir pedwar cam synthesis yn y testun. Mae blociau isoprene mewn squalene wedi'u marcio gan linellau coch wedi'u chwalu.

Cam (1). Synthesis mevalonate o asetad. Mae cam cyntaf biosynthesis colesterol yn arwain at ffurfio cynnyrch canolradd mevalonate (Ffig. 21-34). Mae'r ddau folecwl asetyl CoA yn cyddwyso i roi CoA acetoacetyl, sy'n cyddwyso â'r trydydd moleciwl CoA asetyl i ffurfio cyfansoddyn chwe charbon β-hydroxy-β-methylglutaryl-CoA (HM G -CoA). Mae'r ddau ymateb cyntaf hyn wedi'u cataleiddio thiolase a NM G -CoA synthase, yn y drefn honno. Cytosolig NM G-CoA synthase Mae'r llwybr metabolaidd hwn yn wahanol i'r isoenzyme mitochondrial, sy'n cataleiddio synthesis NM G -CoA yn ystod ffurfio cyrff ceton (gweler Ffig. 17-18).
Ffig. 21-34. Ffurfio mevalonate o asetyl-CoA. Amlygir ffynhonnell mevalonate C-1 a C-2 o asetyl-CoA mewn pinc.

Mae'r trydydd adwaith yn cyfyngu ar gyflymder y broses gyfan. Ynddo, mae NM G -CoA yn cael ei leihau i fevalonate, y mae pob un o'r ddau foleciwl NА D PH yn darparu dau electron ar ei gyfer. HMG-CoA reductase - protein bilen annatod o ER llyfn, mae'n gwasanaethu, fel y gwelwn yn nes ymlaen, fel prif bwynt rheoleiddio llwybr metabolaidd ffurfio colesterol.
Cam (2). Trosi mevalonate yn ddau isoprene actifedig. Yn ystod cam nesaf synthesis colesterol, trosglwyddir tri grŵp ffosffad o foleciwlau ATP i mevalonate (Ffig. 21-35). Mae'r ffosffad sydd wedi'i rwymo i'r grŵp hydrocsyl yn mevalonate C-3 yn y canolradd 3-phospho-5-pyrophosphomevalonate yn grŵp gadael da, yn y cam nesaf mae'r ddau ffosffad hyn a'r grŵp carboxyl cyfagos yn gadael, gan ffurfio bond dwbl yn y cynnyrch pum carbon ∆ 3 -pyrophosphate isopentenyl. Dyma'r cyntaf o ddau isopren actifedig - y prif gyfranogwyr mewn synthesis colesterol. Mae isomeiddio Δ 3 -isopentenylpyrophosphate yn rhoi ail isoprene wedi'i actifadu pyrophosphate dimethylallyl. Mae synthesis pyrophosphate isopentenyl yng nghytoplasm celloedd planhigion yn digwydd yn ôl y llwybr a ddisgrifir yma. Fodd bynnag, mae cloroplastau planhigion a llawer o facteria yn defnyddio llwybr sy'n annibynnol ar mevalonate. Nid yw'r llwybr amgen hwn i'w gael mewn anifeiliaid, felly mae'n ddeniadol wrth greu gwrthfiotigau newydd.
Ffig. 21-35. Trosi mevalonate yn flociau isoprene actifedig. Mae'r chwe uned actifedig yn cyfuno i ffurfio squalene (gweler Ffigur 21-36). Amlygir y grwpiau gadael o 3-phospho-5-pyrophosphomevalonate mewn pinc. Canolradd damcaniaethol yw cromfachau sgwâr.

Cam (3). Anwedd chwe uned isoprene actifedig i ffurfio squalene. Mae pyrophosphate isopentenyl a phyrophosphate dimethylallyl bellach yn cael cyddwysiad pen i gynffon, lle mae un grŵp pyroffosffad yn symud a chadwyn 10-carbon yn ffurfio. - pyrophosphate geranyl (Ffig. 21-36). (Mae pyrophosphate yn glynu wrth y pen.) Mae pyrophosphate geranyl yn mynd trwy'r cyddwysiad pen-i-gynffon canlynol gyda phyroffosffad isopentenyl, a ffurfiau canolradd 15-carbon. pyrophosphate farnesyl. Yn olaf, mae'r ddau folecwl o farnesyl pyrophosphate yn cyfuno “ben wrth ben”, mae'r ddau grŵp ffosffad yn cael eu tynnu - eu ffurfio squalene.
Ffig. 21-36. Ffurfiant squalene. Mae strwythur squalene sy'n cynnwys 30 o atomau carbon yn digwydd yn ystod cyddwysiadau olynol a actifadir gan flociau isoprene (pum carbon).

Daw'r enwau cyffredin ar gyfer y canolradd hyn o enwau'r ffynonellau y cawsant eu hynysu ohonynt gyntaf. Mae gan Geraniol, cydran o olew rhosyn, arogl geraniwm, ac mae gan farnesol, a geir yn lliwiau'r acacia farnesa, lili o arogl y dyffryn. Mae llawer o arogleuon planhigion naturiol yn perthyn i gyfansoddion sydd wedi'u hadeiladu o flociau isoprene. Mae squalene, sydd wedi'i ynysu gyntaf o afu siarc (rhywogaeth Squalus), yn cynnwys 30 atom carbon: 24 atom yn y brif gadwyn a chwe atom yn yr eilyddion metel.
Cam (4). Trawsnewid squalene yn bedair cylch o gnewyllyn steroid. Yn ffig. 21-37 gwelir yn glir bod strwythur y gadwyn squalene, a sterolau - cylchol. Mae gan bob sterol bedair cylch cyddwys sy'n ffurfio'r niwclews steroid, ac mae pob un ohonynt yn alcoholau â grŵp hydrocsyl yn yr atom C-3, a dyna'r enw Saesneg sterol. O dan weithredu squalene monooxygenase ychwanegir un atom ocsigen o O at ddiwedd y gadwyn squalene 2 a ffurfir epocsid. Mae'r ensym hwn yn ocsidas swyddogaeth gymysg arall (ychwanegwch. 21-1), mae NADPH yn lleihau atom ocsigen arall o O. 2 i H.2 O. Clymiadau Dwbl Cynnyrch squalene-2,3-epocsid wedi'i drefnu fel y gall adwaith rhyfeddol o gyson droi cadwyn o epocsid squalene yn strwythur cylchol. Mewn celloedd anifeiliaid, mae'r cylchrediad hwn yn arwain at ffurfio lanosterol sy'n cynnwys pedair cylch sy'n nodweddiadol o'r niwclews steroid. O ganlyniad, mae lanosterol yn cael ei drawsnewid yn golesterol trwy gyfres o oddeutu 20 o ymatebion, sy'n cynnwys ymfudiad rhai grwpiau metel a chael gwared ar eraill. Gwnaethpwyd y disgrifiad o'r llwybr anhygoel hwn o biosynthesis, un o'r rhai anoddaf ymhlith y rhai hysbys, gan Conrad Bloch, Theodore Linen, John Cornfort a George Popiak ddiwedd y 1950au.
Ffig. 21-37. Mae cau cylch yn troi squalene llinol yn graidd steroid cyddwys. Mae'r cam cyntaf yn cael ei gataleiddio gan ocsidas â swyddogaeth gymysg (monooxygenase), a'i cosubstrate yw N AD PH. Mae'r cynnyrch yn epocsid, sydd yn y cam nesaf yn beicio i ffurfio craidd steroid. Cynnyrch terfynol yr adweithiau hyn mewn celloedd anifeiliaid yw colesterol; mewn organebau eraill mae sterolau ychydig yn wahanol iddo yn cael eu ffurfio.


Mae colesterol yn nodwedd sterol o gelloedd anifeiliaid, mae planhigion, ffyngau a phrotestwyr yn cynhyrchu sterolau tebyg iawn eraill.
Maent yn defnyddio'r un llwybr synthesis i squalene-2,3-epoxide, ond yna mae'r llwybrau'n dargyfeirio ychydig, a ffurfir sterolau eraill, fel sigmasterol mewn llawer o blanhigion ac ergosterol mewn ffyngau (Ffig. 21-37).
Enghraifft 21-1 Costau Ynni ar gyfer Synthesis Squalene
Beth yw'r costau ynni (wedi'u mynegi fel moleciwlau ATP) ar gyfer synthesis un moleciwl squalene?
Datrysiad. Wrth synthesis squalene o asetyl-CoA, dim ond pan fydd mevalonate yn cael ei drawsnewid yn rhagflaenydd isoprene squalene wedi'i actifadu y mae ATP yn cael ei wario. Mae angen chwe moleciwl isoprene actifedig i adeiladu moleciwl squalene, ac mae angen tri moleciwl ATP i gynhyrchu pob moleciwl wedi'i actifadu. Yn gyfan gwbl, mae 18 o foleciwlau ATP yn cael eu gwario ar synthesis un moleciwl squalene.
Cyfansoddion colesterol yn y corff
Mewn fertebratau, mae llawer iawn o golesterol yn cael ei syntheseiddio yn yr afu. Mae peth o'r colesterol a syntheseiddiwyd yno wedi'i ymgorffori ym mhilenni hepatocytes, ond mae'n cael ei allforio yn bennaf yn un o'i dair ffurf: colesterol bustlog (bustl), asidau bustl neu esterau colesterol. Asidau bustl ac mae eu halwynau yn ddeilliadau hydroffilig o golesterol, sy'n cael eu syntheseiddio yn yr afu ac yn cyfrannu at dreuliad lipidau (gweler Ffig. 17-1). Esterau colesterol wedi'i ffurfio yn yr afu trwy weithredu acyl-CoA-cholesterol-acyltransferase (ACAT). Mae'r ensym hwn yn cataleiddio trosglwyddiad gweddillion asid brasterog o coenzyme A i'r grŵp hydrocsyl o golesterol (Ffig. 21-38), gan droi colesterol yn ffurf fwy hydroffobig. Mae esterau colesterol mewn gronynnau lipoprotein cyfrinachol yn cael eu cludo i feinweoedd eraill gan ddefnyddio colesterol neu eu storio yn yr afu.
Ffig. 21-38. Synthesis esterau colesterol. Mae Etherification yn gwneud colesterol yn ffurf hydroffobig hyd yn oed yn fwy ar gyfer storio a chludo.

Mae colesterol yn angenrheidiol ar gyfer holl feinweoedd organeb anifeiliaid sy'n tyfu ar gyfer synthesis pilenni, ac mae rhai organau (er enghraifft, y chwarennau adrenal a'r chwarennau rhyw) yn defnyddio colesterol fel rhagflaenydd i hormonau steroid (trafodir hyn isod). Mae colesterol hefyd yn rhagflaenydd i fitamin D (gweler Ffigur 10-20, v. 1).
Mae colesterol a lipidau eraill yn cario lipoproteinau plasma
Mae esterau colesterol a cholesterol, fel triacylglycerols a ffosffolipidau, yn ymarferol anhydawdd mewn dŵr, fodd bynnag, rhaid iddynt symud o'r meinwe y cawsant eu syntheseiddio ynddo i'r meinweoedd lle byddant yn cael eu storio neu eu bwyta. Fe'u cludir gan y llif gwaed ar ffurf lipoproteinau plasma gwaed - cyfadeiladau macromoleciwlaidd proteinau cludwr penodol (apolipoproteinau) gyda ffosffolipidau, colesterol, esterau colesterol a triacylglycerols yn bresennol yn y cyfadeiladau hyn mewn amryw gyfuniadau.
Mae apolipoproteinau (“apo” yn cyfeirio at y protein heb lipid ei hun) yn cyfuno â lipidau i ffurfio sawl ffracsiynau o ronynnau lipoprotein - cyfadeiladau sfferig â lipidau hydroffobig yn y canol a chadwyni asid amino hydroffilig ar yr wyneb (Ffig. 21-39, a). Gyda chyfuniadau amrywiol o lipidau a phroteinau, mae gronynnau o wahanol ddwyseddau yn cael eu ffurfio - o chylomicronau i lipoproteinau dwysedd uchel. Gellir gwahanu'r gronynnau hyn trwy uwchddwysoli (Tabl 21-1) a'u harsylwi'n weledol gan ddefnyddio microsgopeg electron (Ffigur 21-39, b). Mae pob ffracsiwn o lipoproteinau yn cyflawni swyddogaeth benodol, sy'n cael ei bennu gan le synthesis, cyfansoddiad lipid a chynnwys apolipoprotein. Darganfuwyd o leiaf 10 apolipoproteinau gwahanol mewn plasma gwaed dynol (Tabl 21-2), sy'n amrywio o ran maint, adweithiau â gwrthgyrff penodol, a'r dosbarthiad nodweddiadol mewn gwahanol ddosbarthiadau o lipoproteinau. Mae'r cydrannau protein hyn yn gweithredu fel sylweddau signalau sy'n cyfeirio lipoproteinau i feinweoedd penodol neu'n actifadu ensymau sy'n gweithredu ar lipoproteinau.
Tabl 21-1. Lipoproteinau plasma dynol
Cyfansoddiad (ffracsiwn màs,%)
r = 513,000). Mae gronyn o LDL yn cynnwys craidd o oddeutu 1,500 o foleciwlau o esterau colesterol, o amgylch y craidd mae cragen o 500 moleciwl o golesterol, 800 moleciwl o ffosffolipidau ac un moleciwl o apoB-100. b - pedwar dosbarth o lipoproteinau, i'w gweld gyda microsgop electron (ar ôl yr amlygiad o negyddol). Clocwedd, gan ddechrau o'r ffigur chwith uchaf: chylomicrons - gyda diamedr o 50 i 200 nm, PL O NP - o 28 i 70 nm, HDL - o 8 i 11 nm, a LDL - o 20 i 55 nm. Rhoddir priodweddau lipoproteinau yn y tabl. 21-2.

Chylomicrons, y cyfeirir ato yn Sec. 17, symud triacylglycerols bwyd o'r coluddyn i feinweoedd eraill. Dyma'r lipoproteinau mwyaf, mae ganddyn nhw'r dwysedd isaf a'r cynnwys cymharol uchaf o triacylglycerols (gweler Ffig. 17-2). Mae chylomicronau yn cael eu syntheseiddio yn yr ER o gelloedd epithelial sy'n leinio'r coluddyn bach, yna'n symud trwy'r system lymffatig ac yn mynd i mewn i'r llif gwaed trwy'r wythïen is-ddosbarth chwith. Mae apolipoproteinau Chylomicron yn cynnwys apoB-48 (unigryw ar gyfer y dosbarth hwn o lipoproteinau), apoE ac apoC-II (Tabl 21-2). Mae AroC-II yn actifadu lipas lipoprotein yng nghapilarïau meinwe adipose, y galon, cyhyrau ysgerbydol a chwarren mamari sy'n llaetha, gan sicrhau llif asidau brasterog rhydd i'r meinweoedd hyn. Felly, mae chylomicrons yn trosglwyddo asidau brasterog bwyd i feinweoedd, lle byddant yn cael eu bwyta neu eu storio fel tanwydd (Ffig. 21-40). Mae gweddillion chylomicron (wedi'u rhyddhau'n bennaf o triacylglycerols, ond sy'n dal i gynnwys colesterol, apoE ac apoB-48) yn cael eu cludo gan y llif gwaed i'r afu. Yn yr afu, mae derbynyddion yn rhwymo i apoE sydd wedi'i gynnwys mewn gweddillion chylomicron ac yn cyfryngu eu hamsugno gan endocytosis. Mewn hepatocytes, mae'r gweddillion hyn yn rhyddhau eu colesterol ac yn cael eu dinistrio mewn lysosomau.
Tabl 21-2. Apolipoproteinau plasma lipoprotein dynol
Swyddogaeth (os yw'n hysbys)
Yn actifadu L CAT, yn rhyngweithio â chludwr ABC
Yn atal L CAT
Yn actifadu L CAT, cludo / clirio colesterol
Yn rhwymo i'r derbynnydd LDL
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Yn dechrau clirio gweddillion VLDL a chylomicron
Pan fydd bwyd yn cynnwys mwy o asidau brasterog nag y gellir ei ddefnyddio fel tanwydd ar hyn o bryd, maent yn troi'n triacylglycerols yn yr afu, sy'n ffurfio ffracsiwn ag apolipoproteinau penodol lipoproteinau dwysedd isel iawn (VLDL). Gellir trosi carbohydradau gormodol yn yr afu hefyd i triacylglycerols a'u hallforio fel VLDL (Ffig. 21-40, a).Yn ogystal â triacylglycerols, mae'r ffracsiwn VLDL yn cynnwys rhywfaint o esterau colesterol a cholesterol, yn ogystal ag apoB-100, apoC-1, apoC-II, apoC III ac apoE (Tabl 21-2). Mae'r lipoproteinau hyn hefyd yn cael eu cludo gan waed o'r afu i feinwe cyhyrau ac adipose, lle, ar ôl i lipoprotein lipase gael ei actifadu gan apo-C II, mae asidau brasterog am ddim yn cael eu rhyddhau o triacylglycerols o'r ffracsiwn VLDL. Mae adipocytes yn dal asidau brasterog am ddim, unwaith eto yn eu troi'n triacylglycerols, sy'n cael eu storio yn y celloedd hyn ar ffurf cynhwysiant lipid (diferion), i'r gwrthwyneb, yn ocsideiddio asidau brasterog ar unwaith er mwyn cynhyrchu egni. Mae'r rhan fwyaf o weddillion VLDL yn cael eu dileu o'r cylchrediad gan hepatocytes. Mae eu hamsugno, yn debyg i amsugno chylomicronau, yn cael ei gyfryngu gan dderbynyddion ac mae'n dibynnu ar bresenoldeb apoE yng ngweddillion VLDL (yn ychwanegiad 21-2 disgrifir y berthynas rhwng apoE a chlefyd Alzheimer).
Ffig. 21-40. Mae lipoproteinau a chludiant lipid, a - lipidau yn cael eu cludo gan y llif gwaed ar ffurf lipoproteinau, sy'n cael eu cyfuno i sawl ffracsiynau â gwahanol swyddogaethau a chyfansoddiad gwahanol proteinau a lipidau (tab. 21-1, 21-2) ac mae'n cyfateb i ddwysedd y ffracsiynau hyn. Mae lipidau bwyd yn cael eu crynhoi i mewn i chylomicrons, mae'r rhan fwyaf o'r triacylglycerols sydd ynddynt yn cael eu rhyddhau gan lipoprotein lipase i mewn i adipose a meinwe cyhyrau yn y capilarïau. Mae gweddillion chylomicron (sy'n cynnwys protein a cholesterol yn bennaf) yn cael eu dal gan hepatocytes. Mae lipidau mewndarddol a cholesterol o'r afu yn cael eu danfon i adipose a meinwe cyhyrau ar ffurf VLDL. Mae rhyddhau lipidau o VLDL (ynghyd â cholli rhai apolipoproteinau) yn trosi VLDLP yn LDL yn raddol, sy'n dosbarthu colesterol i feinweoedd allhepatig neu'n ei ddychwelyd i'r afu. Mae'r afu yn dal gweddillion VLDL, LDL a gweddillion chylomicronau gan endocytosis wedi'i gyfryngu â derbynnydd. Mae colesterol gormodol mewn meinweoedd allhepatig yn cael ei gludo yn ôl i'r afu ar ffurf LDL. Yn yr afu, mae rhan o'r colesterol yn troi'n halwynau bustl. b - samplau plasma gwaed a gymerwyd ar ôl llwgu (chwith) ac ar ôl bwyta bwyd â chynnwys braster uchel (dde). Mae chylomicronau a ffurfir trwy fwyta bwydydd brasterog yn rhoi tebygrwydd allanol i'r plasma i laeth.

Gyda cholli triacylglycerols, mae cyfran o VLDL yn cael ei throsi i weddillion VLDL, a elwir hefyd yn lipoproteinau dwysedd canolraddol (VLDL), mae tynnu triacylglycerols ymhellach o VLDL yn rhoi lipoproteinau dwysedd isel (LDL) (tab. 21-1). Mae'r ffracsiwn LDL, sy'n gyfoethog iawn mewn esterau colesterol a cholesterol, ac sydd hefyd yn cynnwys apoB-100, yn trosglwyddo colesterol i feinweoedd allhepatig sy'n cario derbynyddion penodol gan gydnabod apoB-100 ar eu pilenni plasma. Mae'r derbynyddion hyn yn cyfryngu'r nifer sy'n cymryd colesterol a esterau colesterol (fel y disgrifir isod).
Ychwanegiad 21-2.Mae alelau ApoE yn pennu nifer yr achosion o glefyd Alzheimer
Yn y boblogaeth ddynol, mae tri amrywiad hysbys (tair alel) o'r genyn sy'n amgodio apolipoprotein E. O'r alelau apoE, yr alele APOEZ yw'r mwyaf cyffredin mewn bodau dynol (tua 78%), yr alelau APOE4 ac APOE2 yw 15 a 7%, yn y drefn honno. Mae alel APOE4 yn arbennig o nodweddiadol o bobl â chlefyd Alzheimer, ac mae'r berthynas hon yn caniatáu rhagweld y bydd y clefyd yn debygol iawn. Mae gan bobl sydd wedi etifeddu APOE4 risg uchel o ddatblygu clefyd Alzheimer hwyr. Mae pobl homosygaidd ar gyfer APOE4 16 gwaith yn fwy tebygol o ddatblygu'r afiechyd, mae oedran cyfartalog y rhai sy'n mynd yn sâl tua 70 oed. I'r bobl sy'n etifeddu dau gopi o AROEZ, i'r gwrthwyneb, mae oedran cyfartalog clefyd Alzheimer yn fwy na 90 mlynedd.
Nid yw'r sylfaen foleciwlaidd ar gyfer y cysylltiad rhwng apoE4 a chlefyd Alzheimer yn hysbys o hyd. Yn ogystal, mae'n dal yn aneglur sut y gall apoE4 effeithio ar dwf cordiau amyloid, sydd, mae'n debyg, yn wraidd y clefyd Alzheimer (gweler Ffig. 4-31, v. 1). Mae rhagdybiaethau'n canolbwyntio ar rôl bosibl apoE wrth sefydlogi strwythur cytoskeleton niwronau. Mae'r proteinau apoE2 ac apoEZ yn rhwymo i nifer o broteinau sy'n gysylltiedig â microtubules niwronau, tra nad yw apoE4 yn rhwymo. Gall hyn gyflymu marwolaeth niwronau. Beth bynnag y gall y mecanwaith hwn fod, mae'r arsylwadau hyn yn rhoi gobaith am ehangu ein dealltwriaeth o swyddogaethau biolegol apolipoproteinau.
Y pedwerydd math o lipoproteinau - lipoproteinau dwysedd uchel (HDL), mae'r ffracsiwn hwn yn cael ei ffurfio yn yr afu a'r coluddyn bach ar ffurf gronynnau bach llawn protein sy'n cynnwys cymharol ychydig o golesterol ac yn hollol rhydd o esterau colesterol (Ffig. 21-40). Mae'r ffracsiwn HDL yn cynnwys apoA-I, apoC-I, apoC-II ac apolipoproteinau eraill (Tabl 21-2), yn ogystal â lecithin-cholesterol-acyltransferase (LC AT), sy'n cataleiddio ffurfio esterau colesterol o lecithin (phosphatidylcholine) a cholesterol (Ffig. 21-41). Mae L CAT ar wyneb gronynnau HDL sydd newydd eu ffurfio yn trosi'r gweddillion coleomicron a ffosffatidylcholine a VLDL yn esterau colesterol, sy'n dechrau ffurfio'r niwclews, gan drawsnewid y gronynnau HDL disgo newydd eu ffurfio yn ronynnau HDL sfferig aeddfed. Yna dychwelir y lipoprotein llawn colesterol hwn i'r afu, lle mae colesterol yn cael ei “ollwng”, mae peth o'r colesterol hwn yn cael ei droi'n halwynau bustl.
Ffig. 21-41. Yr adwaith wedi'i gataleiddio gan lecithin-cholesterol-acyltransferase (L CAT). Mae'r ensym hwn yn bresennol ar wyneb gronynnau HDL ac yn cael ei actifadu gan apoA-1 (cydran o'r ffracsiwn HDL). Mae esterau colesterol yn cronni y tu mewn i'r gronynnau HDL sydd newydd eu ffurfio, gan eu troi'n HDL aeddfed.

Gellir amsugno HDL yn yr afu gan endocytosis wedi'i gyfryngu â derbynnydd, ond mae o leiaf rhywfaint o'r colesterol HDL yn cael ei ddanfon i feinweoedd eraill trwy fecanweithiau eraill. Gall gronynnau HDL rwymo i broteinau derbynnydd SR - BI ar bilen plasma celloedd yr afu ac mewn meinwe steroidogenig fel y chwarennau adrenal. Nid yw'r derbynyddion hyn yn cyfryngu endocytosis, ond trosglwyddiad colesterol a lipidau eraill y ffracsiwn HDL yn rhannol ac yn ddetholus i'r gell. Yna mae'r ffracsiwn HDL “disbyddedig” yn mynd i mewn i'r llif gwaed, lle mae'n cynnwys dognau newydd o lipidau o chylomicronau a gweddillion VLDL. Gall yr un HDL hefyd ddal colesterol sy'n cael ei storio mewn meinweoedd allhepatig a'i drosglwyddo i'r afu gan gwrthdroi cludo colesterol (Ffig. 21-40). Yn un o'r amrywiadau cludo i'r gwrthwyneb, mae rhyngweithiad yr HDL sy'n deillio o hynny â derbynyddion SR-BI mewn celloedd llawn colesterol yn cychwyn trylediad goddefol o golesterol o wyneb y gell i ronynnau HDL, sydd wedyn yn trosglwyddo colesterol yn ôl i'r afu. Mewn amrywiad arall o gludiant i'r gwrthwyneb mewn cell colesterol gyfoethog, ar ôl holltiad HDL, mae apoA-I yn rhyngweithio â'r cludwr gweithredol, protein ABC. Mae ApoA-I (ac yn ôl pob tebyg HDL) yn cael ei amsugno gan endocytosis, yna'n cael ei gyfrinachu eto, wedi'i lwytho â cholesterol, sy'n cael ei gludo i'r afu.
Mae protein ABC1 yn rhan o deulu mawr o gludwyr llawer o gyffuriau, weithiau gelwir y cludwyr hyn yn gludwyr ABC, gan eu bod i gyd yn cynnwys casetiau sy'n rhwymo ATP (ATP - casetiau rhwymo), mae ganddyn nhw hefyd ddau barth traws-bilen gyda chwe heli traws-bilen (gweler caib. . 11, adn. 1). Mae'r proteinau hyn yn trosglwyddo llawer o ïonau, asidau amino, fitaminau, hormonau steroid a halwynau bustl trwy bilenni plasma. Cynrychiolydd arall o'r teulu hwn o gludwyr yw'r protein CFTR, sydd, gyda ffibrosis systig, wedi'i ddifrodi (gweler ychwanegu. 11-3, v. 1).
Mae esterau colesterol yn mynd i mewn i'r gell trwy endocytosis wedi'i gyfryngu â derbynnydd
Mae pob gronyn LDL yn y llif gwaed yn cynnwys apoB-100, sy'n cael ei gydnabod gan broteinau derbynnydd wyneb penodol -Derbynyddion LDL ar bilen y celloedd sydd angen dal colesterol. Mae rhwymo LDL i'r derbynnydd LDL yn cychwyn endocytosis, oherwydd mae LDL a'i dderbynnydd yn symud i'r gell y tu mewn i'r endosome (Ffig. 21-42). Yn y pen draw, mae'r endosome yn asio â'r lysosom, sy'n cynnwys ensymau sy'n hydroli esterau colesterol, gan ryddhau colesterol ac asidau brasterog i'r cytosol. Mae ApoB-100 o LDL hefyd yn torri i lawr i ffurfio asidau amino sy'n cael eu secretu i'r cytosol, ond mae'r derbynnydd LDL yn osgoi diraddio ac yn dychwelyd i wyneb y gell i gymryd rhan eto yn y nifer sy'n derbyn LDL. Mae ApoB-100 hefyd yn bresennol yn VLDL, ond nid yw ei barth rhwymo derbynnydd yn gallu rhwymo i'r derbynnydd LDL; mae trosi VLDLP i LDL yn gwneud y parth rhwymo derbynnydd yn apoB-100 yn hygyrch. Astudiwyd y llwybr cludo colesterol gwaed hwn a'i endocytosis wedi'i gyfryngu â derbynnydd mewn meinweoedd targed gan Michael Brown a Joseph Goldstein.
Michael Brown a Joseph Goldstein

Ffig. 21-42. Dal colesterol gan endocytosis wedi'i gyfryngu â derbynnydd.

Gellir ymgorffori colesterol, sy'n mynd i mewn i'r celloedd fel hyn, mewn pilenni neu ei ail-esterified gan ACAT (Ffig. 21-38) i'w storio yn y cytosol y tu mewn i ddefnynnau lipid. Pan fydd digon o golesterol ar gael yn y ffracsiwn LDL o waed, atalir cronni colesterol mewngellol gormodol trwy leihau cyfradd ei synthesis.
Mae'r derbynnydd LDL hefyd yn rhwymo i apoE ac yn chwarae rhan sylweddol yn y nifer sy'n derbyn chylomicronau a gweddillion VLDL gan yr afu. Fodd bynnag, os nad oes derbynyddion LDL ar gael (fel, er enghraifft, mewn straen llygoden gyda genyn derbynnydd LDL ar goll), mae gweddillion VLDL a chylomicrons yn dal i gael eu hamsugno gan yr afu, er nad yw LDL yn cael ei amsugno. Mae hyn yn dynodi presenoldeb system wrth gefn ategol ar gyfer endocytosis wedi'i gyfryngu â derbynnydd o weddillion VLDL a chylomicron. Un o'r derbynyddion wrth gefn yw'r protein LRP (protein sy'n gysylltiedig â derbynnydd lipoprotein), sy'n gysylltiedig â derbynyddion lipoprotein, sy'n rhwymo i apoE a nifer o ligandau eraill.
Sawl lefel o reoleiddio biosynthesis colesterol
Mae synthesis colesterol yn broses gymhleth ac egnïol o ddrud, felly mae'n amlwg bod y corff yn fuddiol cael mecanwaith ar gyfer rheoleiddio biosynthesis colesterol, sy'n ailgyflenwi ei swm yn ychwanegol at yr hyn sy'n dod gyda bwyd. Mewn mamaliaid, mae cynhyrchu colesterol yn cael ei reoleiddio gan grynodiad mewngellol
glwcagon colesterol a hormonau ac inswlin. Mae cam trosi HMG - CoA i mevalonate (Ffig. 21-34) yn cyfyngu ar gyflymder llwybr metabolig ffurfio colesterol (prif bwynt rheoleiddio). Mae'r adwaith hwn yn cael ei gataleiddio gan HMG - CoA reductase. Mae rheoleiddio mewn ymateb i newidiadau mewn lefelau colesterol yn cael ei gyfryngu gan system reoleiddio trawsgrifio cain ar gyfer genyn sy'n amgodio HMG - CoA reductase. Mae'r genyn hwn, ynghyd â mwy nag 20 o enynnau eraill sy'n amgodio ensymau sy'n ymwneud ag amsugno a synthesis colesterol ac asidau brasterog annirlawn, yn cael ei reoli gan deulu bach o broteinau o'r enw proteinau sy'n rhyngweithio â'r elfen sterol-reoleiddio o ffurfio protein (SREBP, proteinau rhwymo elfen reoleiddio sterol) . Ar ôl synthesis, cyflwynir y proteinau hyn i'r reticulum endoplasmig. Yr unig barth SREBP amino-derfynell hydawdd sy'n gweithredu fel ysgogydd trawsgrifio gan ddefnyddio'r mecanweithiau a ddisgrifir yn Ch. 28 (adn. 3). Fodd bynnag, nid oes gan y parth hwn fynediad i'r niwclews ac ni all gymryd rhan yn actifadu'r genyn cyhyd â'i fod yn aros yn y moleciwl SREBP. Er mwyn actifadu trawsgrifio'r genyn HMG - CoA reductase a genynnau eraill, mae'r parth gweithredol trawsgrifiadol wedi'i wahanu oddi wrth weddill SREBP gan holltiad proteinolytig. Pan fydd colesterol yn uchel, mae proteinau SREBP yn anactif, wedi'u gosod ar ER mewn cymhleth â phrotein arall o'r enw SCAP (SREBP - protein actifadu hollt) (Ffig. 21-43). Mae'n SCAP sy'n clymu colesterol a nifer o sterolau eraill, gan weithredu fel synhwyrydd sterol. Pan fydd lefel y sterol yn uchel, mae'n debyg bod y cymhleth SCAP - SREBP yn rhyngweithio â phrotein arall, sy'n cadw'r cymhleth cyfan yn yr ER. Pan fydd lefel y sterolau yn y gell yn gostwng, mae'r newid cydffurfiol yn SCAP yn arwain at golli gweithgaredd cadw, ac mae'r cymhleth SCAP - SREBP yn mudo y tu mewn i'r fesiglau i gyfadeilad Golgi. Yn y cymhleth Golgi, mae proteinau SREBP yn cael eu clirio ddwywaith gan ddau brotein gwahanol, yr ail holltiad sy'n rhyddhau'r parth amino-derfynell i'r cytosol. Mae'r parth hwn yn symud i'r niwclews ac yn actifadu trawsgrifio genynnau targed. Mae gan barth protein amino-derfynell SREBP hanner oes fer ac mae'n cael ei ddiraddio'n gyflym gan broteasomau (gweler Ffig. 27-48, t. 3). Pan fydd y lefel sterol yn codi'n ddigonol, mae rhyddhau proteinolytig parthau protein SR EBP gyda'r derfynfa amino yn cael ei rwystro eto, ac mae diraddiad proteasome'r parthau gweithredol presennol yn arwain at gau'r genynnau targed yn gyflym.
Ffig. 21-43. Actifadu SR EBP. Mae proteinau SREB P sy'n rhyngweithio ag elfen a reoleiddir gan sterol (lliw gwyrdd), yn syth ar ôl synthesis, yn cael eu cyflwyno i'r ER, gan ffurfio cymhleth â S CAP (lliw coch). (Mae N a C yn dynodi pennau amin a charboxyl y proteinau.) Yn y cyflwr wedi'i rwymo gan S-CAP, mae proteinau SRE BP yn anactif. Pan fydd y lefel sterol yn gostwng, mae cymhleth CAP SR EBP-S yn mudo i gyfadeilad Golgi, ac mae'r proteinau SR EBP yn cael eu clirio yn olynol gan ddau brotein gwahanol. Mae'r parth protein SR EBP terfynell asid amino rhydd yn mudo i'r niwclews, lle mae'n actifadu trawsgrifio genynnau a reoleiddir gan sterol.

Mae synthesis colesterol hefyd yn cael ei reoleiddio gan sawl mecanwaith arall (Ffig. 21-44). Mae rheolaeth hormonaidd yn cael ei gyfryngu gan addasiad cofalent NM G-CoA reductase. Mae'r ensym hwn yn bodoli mewn ffurfiau ffosfforyleiddiedig (anactif) a dadffosfforyleiddiedig (gweithredol). Mae glwcagon yn ysgogi ffosfforyleiddiad (anactifadu) yr ensym, ac mae inswlin yn hyrwyddo dadffosfforyleiddiad, gan actifadu'r ensym ac yn ffafrio synthesis colesterol. Mae crynodiadau mewngellol uchel o golesterol yn actifadu ASAT, sy'n cynyddu esterification colesterol i'w ddyddodi. Yn olaf, mae lefelau uchel o golesterol cellog yn rhwystro trawsgrifio genyn sy'n amgodio derbynnydd LDL, gan leihau cynhyrchiant y derbynnydd hwn ac, felly, y nifer sy'n cymryd colesterol o'r gwaed.
Ffig. 21-44. Mae rheoleiddio lefelau colesterol yn darparu cydbwysedd rhwng synthesis ac amsugno colesterol o fwyd. Mae glwcagon yn hwyluso ffosfforyleiddiad (anactifadu) NM G -CoA reductase, mae inswlin yn hyrwyddo dadffosfforyleiddiad (actifadu). X - metabolion colesterol anhysbys sy'n ysgogi proteolysis NM G -CoA reductase.

Gall colesterol heb ei reoleiddio arwain at salwch difrifol mewn pobl. Pan fydd cyfanswm y colesterol synthetig a'r colesterol a geir o fwyd yn fwy na'r swm sydd ei angen ar gyfer cydosod pilenni, synthesis halwynau bustl a steroidau, gall croniadau patholegol o golesterol mewn pibellau gwaed (placiau atherosglerotig) ymddangos, gan arwain at eu rhwystr (atherosglerosis). Mewn gwledydd diwydiannol, methiant y galon oherwydd rhwystro'r rhydwelïau coronaidd yw prif achos marwolaeth. Mae datblygiad atherosglerosis yn gysylltiedig â lefel uchel o golesterol yn y gwaed ac yn enwedig gyda lefel uchel o golesterol a drosglwyddir gyda'r ffracsiwn LDL, ac mae lefel uchel o waed HDL, i'r gwrthwyneb, yn effeithio'n ffafriol ar gyflwr pibellau gwaed.
Gyda hypercholesterolemia etifeddol (nam genetig), mae lefel colesterol yn y gwaed yn uchel iawn - mae atherosglerosis difrifol yn datblygu yn y bobl hyn sydd eisoes yn ystod plentyndod. Oherwydd derbynnydd LDL diffygiol, nid oes digon o golesterol LDL wedi'i gyfryngu gan y derbynnydd. O ganlyniad, nid yw colesterol yn cael ei dynnu o'r llif gwaed, mae'n cronni ac yn cyfrannu at ffurfio placiau atherosglerotig. Mae'r synthesis o golesterol mewndarddol yn parhau er gwaethaf y colesterol gormodol yn y gwaed, gan na all colesterol allgellog fynd i mewn i'r gell i reoleiddio synthesis mewngellol (Ffig. 21 -44).Ar gyfer trin cleifion â hypercholesterolemia etifeddol a chlefydau eraill sy'n gysylltiedig â cholesterol serwm uchel, defnyddir dosbarthiadau statin. Mae rhai ohonynt ar gael o ffynonellau naturiol, tra bod eraill yn cael eu syntheseiddio gan y diwydiant fferyllol. Mae statinau yn debyg i mevalonate (ychwanegwch. 21-3) ac maent yn atalyddion cystadleuol NMS-CoA reductase.
Ychwanegiad 21-3. MEDDYGINIAETH. Y rhagdybiaeth lipid a chreu statinau
Clefyd coronaidd y galon (CHD) yw prif achos marwolaeth mewn gwledydd datblygedig. Mae culhau'r rhydwelïau coronaidd sy'n cludo gwaed i'r galon yn digwydd o ganlyniad i ffurfio dyddodion brasterog o'r enw placiau atherosglerotig; mae'r placiau hyn yn cynnwys colesterol, proteinau ffibrillar, calsiwm, ceuladau platennau, a darnau celloedd. Yn yr XXfed ganrif. Bu dadl weithredol am y berthynas rhwng rhwystro prifwythiennol (atherosglerosis) a cholesterol yn y gwaed. Mae'r trafodaethau a'r ymchwil weithredol i'r cyfeiriad hwn wedi arwain at greu cyffuriau effeithiol sy'n gostwng colesterol.
Ym 1913, cyhoeddodd N.N. Anichkov, gwyddonydd ac arbenigwr adnabyddus o Rwsia ym maes patholeg arbrofol, waith lle dangosodd fod cwningod sy'n cael eu bwydo â bwyd sy'n llawn colesterol yn datblygu niwed i bibellau gwaed sy'n debyg i blaciau atherosglerotig yn llestri pobl oedrannus. Cynhaliodd Anichkov ei ymchwil am sawl degawd a chyhoeddodd y canlyniadau mewn cyfnodolion adnabyddus o'r Gorllewin. Yn anffodus, ni ddaeth ei ddata yn sail i fodel ar gyfer datblygu atherosglerosis mewn pobl, oherwydd ar y pryd roedd y rhagdybiaeth yn bodoli bod y clefyd hwn yn ganlyniad naturiol i heneiddio ac na ellir ei atal. Fodd bynnag, roedd tystiolaeth yn cronni'n raddol o berthynas rhwng colesterol serwm a datblygiad atherosglerosis (rhagdybiaeth lipid), ac yn y 1960au. mae rhai ymchwilwyr wedi nodi'n benodol y gellir trin y clefyd hwn â meddyginiaethau. Fodd bynnag, roedd y safbwynt arall yn bodoli nes cyhoeddi canlyniadau astudiaeth eang o rôl colesterol a gynhaliwyd gan Sefydliad Iechyd Cenedlaethol yr UD (Treial Atal Sylfaenol Coronaidd) ym 1984. Dangoswyd gostyngiad ystadegol arwyddocaol yn amlder cnawdnychiant myocardaidd a strôc gyda gostyngiad mewn colesterol yn y gwaed. Yn yr astudiaeth hon, defnyddiwyd colesterol, resin cyfnewid anion sy'n clymu asidau bustl, i ostwng colesterol. Mae'r canlyniadau wedi ysgogi'r chwilio am gyffuriau therapiwtig newydd, mwy pwerus. Rhaid imi ddweud, yn y byd gwyddonol, bod amheuon ynghylch dilysrwydd y rhagdybiaeth lipid wedi diflannu'n llwyr dim ond gyda dyfodiad statinau ar ddiwedd yr 1980au - dechrau'r 1990au.
Darganfuwyd y statin cyntaf gan Akira Endo yn Sankyo yn Tokyo. Cyhoeddodd Endo ei waith ym 1976, er iddo ddelio â phroblem metaboledd colesterol am sawl blwyddyn. Ym 1971, awgrymodd y gallai atalyddion synthesis colesterol hefyd gael eu cynnwys yn y cynhyrchwyr madarch o wrthfiotigau a astudiwyd bryd hynny. Am sawl blwyddyn o waith dwys, dadansoddodd fwy na 6,000 o ddiwylliannau o wahanol fadarch, nes iddo ddod i ganlyniad cadarnhaol. Enw'r cyfansoddyn o ganlyniad oedd compactin. Gwnaeth y sylwedd hwn ostwng colesterol mewn cŵn a mwncïod. Daliodd yr astudiaethau hyn sylw Michael Brown a Joseph Goldstein o Ysgol Feddygol De-orllewinol Prifysgol Texas. Dechreuodd Brown ac Goldstein, ynghyd ag Endo, astudiaeth ar y cyd a chadarnhaodd ei ddata. Roedd prif lwyddiannau'r treialon clinigol cyntaf yn cynnwys cwmnïau fferyllol yn natblygiad y cyffuriau newydd hyn. Yn Merck, lansiodd tîm dan arweiniad Alfred Alberts a Roy Wagelos ddangosiad newydd o ddiwylliannau madarch ac, o ganlyniad i ddadansoddi cyfanswm o 18 o ddiwylliannau, darganfuwyd cyffur gweithredol arall. Enw'r sylwedd newydd yw lovastatin. Fodd bynnag, ar yr un pryd, credwyd yn eang bod rhoi dosau uchel o gompin i gŵn yn arwain at ddatblygiad canser a chwilio am statinau newydd yn yr 1980au. wedi'i atal. Fodd bynnag, erbyn hynny, roedd buddion defnyddio statinau i drin cleifion â hypercholesterolemia teuluol eisoes yn amlwg. Ar ôl ymgynghori niferus ag arbenigwyr rhyngwladol a'r Weinyddiaeth Bwyd a Chyffuriau (FDA, UDA), dechreuodd Merck ddatblygu lovastatin. Nid yw astudiaethau helaeth dros y ddau ddegawd nesaf wedi datgelu effaith carcinogenig lovastatin a'r genhedlaeth newydd o gyffuriau a ymddangosodd ar ei ôl.
Ffig. 1. Mae statinau yn atalyddion NM G-CoA reductase. Cymhariaeth o strwythur mevalonate a phedwar cynnyrch fferyllol (statinau) sy'n rhwystro gweithred NM G -CoA reductase.

Statinau atal gweithred HMG - CoA - reductase, dynwared strwythur mevalonate, a thrwy hynny rwystro synthesis colesterol. Mewn cleifion â hypercholesterolemia a achosir gan ddiffyg mewn un copi o'r genyn derbynnydd LDL, wrth gymryd lovastatin, mae lefelau colesterol yn cael eu gostwng 30%. Mae'r cyffur hyd yn oed yn fwy effeithiol mewn cyfuniad â resinau arbennig sy'n clymu asidau bustl ac yn atal eu hamsugno o'r coluddion.
Ar hyn o bryd, defnyddir statinau amlaf i ostwng colesterol plasma gwaed. Wrth gymryd unrhyw feddyginiaeth, mae'r cwestiwn yn codi am eu sgîl-effeithiau annymunol. Fodd bynnag, yn achos statinau, mae llawer o sgîl-effeithiau, mewn cyferbyniad, yn gadarnhaol. Gall y cyffuriau hyn ysgogi llif y gwaed, trwsio placiau atherosglerotig sydd eisoes yn bodoli (fel nad ydynt yn torri i ffwrdd o waliau pibellau gwaed ac nad ydynt yn ymyrryd â llif y gwaed), yn atal agregu platennau, a hefyd yn gwanhau prosesau llidiol yn waliau pibellau gwaed. Mewn cleifion sy'n cymryd statinau am y tro cyntaf, mae'r effeithiau hyn yn cael eu hamlygu hyd yn oed cyn i lefelau colesterol ddechrau dirywio, ac o bosibl maent yn gysylltiedig â gwahardd synthesis isoprenoid. Wrth gwrs, nid yw pob sgil-effaith statinau yn fuddiol. Mewn rhai cleifion (fel arfer ymhlith y rhai sy'n cymryd statinau mewn cyfuniad â chyffuriau eraill sy'n gostwng colesterol), gall poen cyhyrau a gwendid cyhyrau ddigwydd, ac weithiau ar ffurf eithaf cryf. Mae sgîl-effeithiau eithaf niferus statinau eraill wedi'u cofrestru hefyd, sydd, yn ffodus, yn anaml yn digwydd. Yn y mwyafrif llethol o gleifion, gall cymryd statinau rwystro datblygiad clefyd cardiofasgwlaidd. Fel unrhyw feddyginiaeth arall, dim ond fel yr argymhellwyd gan eich meddyg y dylid defnyddio statinau.

Gydag absenoldeb etifeddol o golesterol HDL, mae lefelau colesterol yn isel iawn, gyda chlefyd Tangier, yn ymarferol nid yw colesterol yn cael ei bennu. Mae'r ddau anhwylder genetig yn deillio o dreigladau mewn protein ABC1. Ni all y ffracsiwn colesterol heb HDL ddal colesterol o gelloedd diffygiol ABC1, ac mae celloedd sy'n disbyddu colesterol yn cael eu tynnu o'r gwaed yn gyflym a'u dinistrio. Mae absenoldeb etifeddol HDL a chlefyd Tangier yn brin iawn (mae llai na 100 o deuluoedd â chlefyd Tangier yn hysbys ledled y byd), ond mae'r afiechydon hyn yn dangos rôl protein ABC1 wrth reoleiddio lefelau plasma HDL. Gan fod lefelau HDL plasma isel yn cydberthyn â chyfradd uchel o ddifrod rhydwelïau coronaidd, gall protein ABC1 fod yn darged defnyddiol ar gyfer cyffuriau sydd wedi'u cynllunio i reoleiddio lefelau HDL. ■
Mae hormonau steroid yn cael eu ffurfio trwy rannu'r gadwyn ochr o golesterol a'i ocsidiad.
Mae person yn derbyn ei holl hormonau steroid o golesterol (Ffig. 21-45). Mae dau ddosbarth o hormonau steroid yn cael eu syntheseiddio yn y cortecs adrenal: mineralcorticoidau,sy'n rheoleiddio amsugno ïonau anorganig (Na +, C l - a HC O. 3 -) yn yr arennau, a glucocorticoidau, sy'n helpu i reoleiddio gluconeogenesis a lleihau'r ymateb llidiol. Cynhyrchir hormonau rhyw yng nghelloedd atgenhedlu dynion a menywod ac yn y brych. Yn eu plith progesteron sy'n rheoleiddio'r cylch atgenhedlu benywaidd, androgenau (e.e. testosteron) a estrogens (estradiol), sy'n effeithio ar ddatblygiad nodweddion rhywiol eilaidd mewn dynion a menywod, yn y drefn honno. Mae hormonau steroid yn cael effaith ar grynodiadau isel iawn ac felly maent yn cael eu syntheseiddio mewn symiau cymharol fach. O'i gymharu â halwynau bustl, cymharol ychydig o golesterol sy'n cael ei fwyta i gynhyrchu hormonau steroid.

Ffig. 21-45. Mae rhai hormonau steroid yn cael eu ffurfio o golesterol. Dangosir strwythurau rhai o'r cyfansoddion hyn yn Ffig. 10-19, adn. 1.

Mae synthesis hormonau steroid yn gofyn am gael gwared â nifer neu'r cyfan o'r atomau carbon yn “cadwyn ochr” cylch-D C-17 colesterol. Mae tynnu cadwyn ochr yn digwydd ym mitocondria meinweoedd steroidogenig. Mae'r broses dynnu yn cynnwys hydroxylation dau atom carbon cyfagos o'r gadwyn ochr (C-20 a C-22), yna holltiad y bond rhyngddynt (Ffig. 21-46). Mae ffurfio hormonau amrywiol hefyd yn cynnwys cyflwyno atomau ocsigen. Mae'r holl adweithiau hydrocsiad ac ocsidiad yn ystod biosynthesis steroid yn cael eu cataleiddio gan ocsidiadau swyddogaeth gymysg (ychwanegwch 21-1) sy'n defnyddio NА D PH, O 2 a cytochrome mitochondrial P-450.
Ffig. 21-46. Holltiad y gadwyn ochr wrth synthesis hormonau steroid. Yn y system ocsidas hon sydd â swyddogaeth gymysg sy'n ocsideiddio atomau carbon cyfagos, mae cytocrom P-450 yn gweithredu fel cludwr electronau. Hefyd yn rhan o'r broses mae proteinau sy'n cludo electronau, adrenodoxin ac adrenodoxin reductase. Darganfuwyd y system hon o hollti cadwyn ochr ym mitocondria'r cortecs adrenal, lle mae steroidau'n cael eu cynhyrchu'n weithredol. Mae Pregnenolone yn rhagflaenydd i'r holl hormonau steroid eraill (Ffig. 21-45).

Mae canolradd biosynthesis colesterol yn cymryd rhan mewn llawer o lwybrau metabolaidd eraill.
Yn ychwanegol at ei rôl fel canolradd o biosynthesis colesterol, mae pyrophosphate isopentenyl yn gwasanaethu fel rhagflaenydd wedi'i actifadu wrth synthesis nifer enfawr o fiomoleciwlau sy'n cyflawni swyddogaethau biolegol amrywiol (Ffig. 21-47). Mae'r rhain yn cynnwys fitaminau A, E a K, pigmentau planhigion fel caroten a'r gadwyn ffytol cloroffyl, rwber naturiol, llawer o olewau hanfodol (er enghraifft, sylfaen persawrus olew lemwn, ewcalyptws, mwsg), hormon ifanc pryfed sy'n rheoleiddio metamorffosis, dolicholau, sy'n gwasanaethu fel cludwyr toddadwy lipid wrth synthesis cymhleth polysacaridau, ubiquinone a plastoquinone - cludwyr electronau mewn mitocondria a chloroplastau. Mae'r holl foleciwlau hyn yn isoprenoidau o ran strwythur. Mae mwy na 20,000 o wahanol isoprenoidau wedi'u darganfod ym myd natur, ac mae cannoedd o rai newydd yn cael eu riportio bob blwyddyn.
Ffig. 21-47. Y darlun cyffredinol o biosynthesis isoprenoidau. Rhoddir strwythurau mwyafrif y cynhyrchion terfynol a gyflwynir yma ym mhen. 10 (adn. 1).

Mae Prenylation (atodiad cofalent isoprenoid, gweler Ffig. 27-35) yn fecanwaith cyffredin lle mae proteinau'n angori ar wyneb mewnol pilenni celloedd mamalaidd (gweler Ffig. 11-14). Mewn rhai proteinau, mae'r lipid wedi'i rwymo yn cael ei gynrychioli gan grŵp barnesyl 15-carbon, mewn eraill mae'n grŵp geranyl geranyl 20-carbon. Mae'r ddau fath hyn o lipidau yn atodi gwahanol ensymau. Mae'n bosibl bod adweithiau prenylation yn cyfeirio proteinau i wahanol bilenni yn dibynnu ar ba lipid sydd ynghlwm. Mae lledaenu protein yn rôl bwysig arall ar gyfer deilliadau isoprene - cyfranogwyr y llwybr metabolaidd colesterol.
Crynodeb o Adran 21.4 Biosynthesis Colesterol, Steroidau ac Isoprenoidau
■ Mae colesterol yn cael ei ffurfio o asetyl-CoA mewn dilyniant adwaith cymhleth trwy gyfryngol fel β-hydroxy-β-methylglutaryl-CoA, mevalonate, dau pyrophosphate dimethylallyl isoprene actifedig a pyrophosphate isopentenyl. Mae cyddwysiad unedau isoprene yn rhoi squalene nad yw'n gylchol, sy'n beicio i ffurfio system gylch gyddwys a chadwyn ochr steroid.
■ Mae synthesis colesterol o dan reolaeth hormonaidd ac, ar ben hynny, mae'n cael ei rwystro gan grynodiadau cynyddol o golesterol mewngellol, sy'n digwydd trwy addasu cofalent a rheoleiddio trawsgrifio.
■ Mae'r gwaed yn cario esterau colesterol a cholesterol fel lipoproteinau plasma. Mae'r ffracsiwn VLDL yn trosglwyddo colesterol, esterau colesterol a triacylglycerols o'r afu i feinweoedd eraill, lle mae'r triacylglycerols yn cael ei glirio gan lipas lipoprotein a VLDL yn cael ei drawsnewid yn LDL. Mae'r ffracsiwn LDL sydd wedi'i gyfoethogi mewn esterau colesterol a cholesterol yn cael ei ddal yn anuniongyrchol gan dderbynyddion gan endocytosis, tra bod apolipoprotein B-100 mewn LDL yn cael ei gydnabod gan dderbynyddion pilen plasma. Mae HDL yn tynnu colesterol o'r gwaed, gan ei drosglwyddo i'r afu. Gall cyflyrau maethol neu ddiffygion genetig mewn metaboledd colesterol arwain at atherosglerosis a cnawdnychiant myocardaidd.
■ Mae hormonau steroid (glucocorticoids, mineralocorticoids a hormonau rhyw) yn cael eu ffurfio o golesterol trwy newid y gadwyn ochr a chyflwyno atomau ocsigen i system steroid y cylchoedd. Mae llawer o gyfansoddion isoprenoid eraill yn cael eu cynhyrchu o fevalonate trwy gyddwysiad pyrophosphate isopentenyl a phyroffosffad dimethylallyl ynghyd â cholesterol.
■ Mae Prenylation rhai proteinau yn eu cyfeirio at wefannau rhwymol â philenni celloedd ac mae'n bwysig ar gyfer eu gweithgaredd biolegol.
Cwestiwn 48. Rheoleiddio metaboledd asidau brasterog uchel (β-ocsidiad a biosynthesis). Synthesis o CoA malonyl. Carboxylase Acetyl CoA, rheoleiddio ei weithgaredd. Cludo acyl Co-a trwy bilen fewnol mitocondria.
Prif
faint o ffenylalanîn sy'n cael ei fwyta
mewn 2 ffordd:
yn troi ymlaen
mewn gwiwerod,
troi
mewn tyrosine.
Troi
phenylalanine i tyrosine yn bennaf
angenrheidiol i gael gwared ar ormodedd
ffenylalanîn, ers crynodiadau uchel
Mae'n wenwynig i gelloedd. Addysg
does dim ots tyrosine mewn gwirionedd
ers diffyg yr asid amino hwn
mewn celloedd yn ymarferol nid yw'n digwydd.
Prif
mae metaboledd ffenylalanîn yn dechrau
gyda'i hydroxylation (Ffig. 9-29), yn
gan arwain at tyrosine.
Mae'r adwaith hwn yn cael ei gataleiddio gan benodol
monooxy-nase - hydra phenylalanine (zsilase,
sy'n gwasanaethu fel cyd-gynhyrchydd
tetrahydrobiopterin (N4BP).
Mae gweithgaredd ensym hefyd yn dibynnu ar
presenoldeb Fe2.
Yn
mae'r afu yn cael ei gyflymu yn bennaf
glycogen (gweler adran 7). Fodd bynnag stociau
mae glycogen yn yr afu wedi'i ddisbyddu
18-24 awr o ymprydio. Prif ffynhonnell
glwcos wrth i'r stociau ddod i ben
daw glycogen yn gluconeogenesis,
sy'n dechrau cyflymu drwodd
Ffig.
11-29. Newidiadau metabolaidd mawr
egni wrth newid amsugnol
cyflwr postabsorbent. CT
- cyrff ceton, FA - asidau brasterog.
4-6 h
ar ôl y pryd olaf. Is-haenau
defnyddir glyserol ar gyfer synthesis glwcos,
asidau amino a lactad. Yn uchel
cyfradd synthesis crynodiad glwcagon
asidau brasterog yn cael eu lleihau oherwydd
ffosfforyleiddiad ac anactifadu
carboxylase a chyfradd asetyl CoA
mae p-ocsidiad yn cynyddu. Fodd bynnag,
mwy o gyflenwad braster i'r afu
asidau sy'n cael eu cludo
o ddepos braster. Ffurfiwyd asetyl-CoA
wrth ocsideiddio asidau brasterog, fe'i defnyddir
yn yr afu ar gyfer synthesis cyrff ceton.
Yn
meinwe adipose gyda chrynodiad cynyddol
cyfradd synthesis llai glwcagon
Mae TAG a lipolysis yn cael ei ysgogi. Ysgogi
lipolysis - canlyniad actifadu
lipase TAG sy'n sensitif i hormonau
adipocytes dan ddylanwad glwcagon.
Daw Asidau Brasterog yn Bwysig
ffynonellau egni yn yr afu, y cyhyrau a
meinwe adipose.
Felly
felly, yn y cyfnod postabsorption
cynhelir crynodiad glwcos yn y gwaed
ar y lefel o 80-100 mg / dl, a lefel y brasterog
cyrff asidau a ceton yn cynyddu.
Siwgr
mae diabetes yn glefyd sy'n digwydd
oherwydd absoliwt neu gymharol
diffyg inswlin.
A.
Y prif ffurfiau clinigol o siwgr
diabetes
Yn ôl
Sefydliad y Byd
diabetes gofal iechyd
dosbarthu yn ôl gwahaniaethau
ffactorau genetig a chlinigol
dwy brif ffurf: diabetes
Math I - dibynnol ar inswlin (IDDM), a diabetes
Math II - annibynnol nad yw'n inswlin (NIDDM).
Rheoliad
synthesis o ensym rheoleiddio zhk
synthesis o lcd - carboxylase CoA asetyl.
Mae'r ensym hwn yn cael ei reoleiddio gan sawl un
ffyrdd.
Actifadu / Dadgysylltiad
cyfadeiladau subunit ensymau. Yn
ffurf anactif o carboxylase asetyl CoA
yn cynrychioli cyfadeiladau ar wahân,
mae pob un yn cynnwys 4 is-uned.
Mae actifadydd yr ensym yn sitrad. Mae'n ysgogi
cyfuniad o gyfadeiladau, o ganlyniad
lle mae gweithgaredd ensymau yn cynyddu
. Atalydd-palmitoyl-CoA. Mae'n galw
daduniad a gostyngiad cymhleth
gweithgaredd ensymau.
Ffosfforyleiddiad / Deffosfforyleiddiad
carboxylase asetyl CoA. Yn
cyflwr postabsorption neu mewn
gwaith corfforol wedi'i glwcagonoli
adrenalin trwy gyclase adenylate
gweithredir y system gan prokinase A a
ysgogi ffosfforyleiddiad subunit
carboxylase asetyl CoA. Ffosfforyleiddiedig
mae'r ensym yn anactif ac yn synthesis brasterog
asidau yn stopio.
Amsugnol
inswlin cyfnod yn actifadu ffosffatase,
ac mae carboxylase asetyl-CoA yn mynd i mewn
cyflwr dephosphorylated. Yna
o dan ddylanwad sitrad yn digwydd
polymerization protomers yr ensym, a
mae'n dod yn egnïol. Yn ogystal ag actifadu
ensym, sitrad yn perfformio un arall
swyddogaeth wrth synthesis LCD. Amsugnol
cyfnod ym mitocondria celloedd yr afu
yn cronni sitrad, lle mae
cludir y gweddillion acyl i
cytosol.
Rheoliad
cyfraddau β-ocsidiad.
Llwybr Β-ocsidiad-metabolig,
wedi'i gysylltu'n gadarn â gwaith y CPE ac yn gyffredinol
ffyrdd o cataboliaeth. Felly ei gyflymder
wedi'i reoleiddio gan angen celloedd am
egni h.y. yn ôl cymarebau ATP / ADP a NADH / NAD, yn ogystal â chyfradd adweithio CPE a
llwybr cyffredin cataboliaeth. Cyflymder
Mae β-ocsidiad mewn meinweoedd yn dibynnu ar argaeledd
swbstrad, h.y.
ar faint o fraster
asidau sy'n mynd i mewn i'r mitocondria.
Crynodiad Asid Brasterog Am Ddim
yn y gwaed yn codi wrth actifadu
lipolysis mewn meinwe adipose yn ystod ymprydio
dan ddylanwad glwcagon ac yn ystod corfforol
gweithio dan ddylanwad adrenalin. Yn y rhain
daw asidau brasterog
prif ffynhonnell egni
ar gyfer cyhyrau ac afu, o ganlyniad i
Mae β-ocsidiadau yn cael eu ffurfio gan NADH ac mae asetyl-CoA yn atal
pyruvate dehydrogenase cymhleth.
Trawsnewid ffurfiad pyruvate
o glwcos i asetyl-CoA yn arafu.
Mae metabolion canolradd yn cronni
glycolysis ac, yn benodol, glwcos-6-ffosffad.
Mae glwcos-6-ffosffad yn atal hexokinase
ac felly yn digalonni
defnyddio glwcos yn y broses
glycolysis. Felly, y trechaf
defnyddio lcd fel y brif ffynhonnell
egni mewn meinwe cyhyrau a'r afu
yn arbed glwcos ar gyfer meinwe nerfau a
celloedd gwaed coch.
Cyfradd ocsidiad ocsidiad hefyd
yn dibynnu ar weithgaredd ensymau
carnitine acyltransferases I.
Yn yr afu, mae'r ensym hwn wedi'i atal.
malonyl CoA, sylwedd a ffurfiwyd
gyda biosynthesis o lcd. Yn y cyfnod amsugnol
mae glycolysis yn cael ei actifadu yn yr afu a
mae ffurfio asetyl-CoA yn cynyddu
o pyruvate. Adwaith synthesis cyntaf
trosi lcd o asetyl-CoA yn malonyl-CoA.
Mae Malonyl-CoA yn atal β-ocsidiad lcd,
y gellir ei ddefnyddio ar gyfer synthesis
braster.
Addysg
malonyl-CoA o asetyl-CoA-rheoliadol
adwaith mewn biosynthesis lcd. Ymateb cyntaf
trosi synthesis lcd o asetyl-CoA i
CoA malonyl. Ensym catalytig
yr adwaith hwn (asetyl Coa carboxylase),
yn perthyn i'r dosbarth o ligases. Mae'n cynnwys
biotin wedi'i rwymo'n gofalent. Yn y cyntaf
camau ymateb cofalent co2
yn rhwymo i biotin oherwydd egni
ATP, yng ngham 2 COO- trosglwyddwyd
ar asetyl-CoA i ffurfio malonyl-CoA.
Gweithgaredd Ensym Carboxylase Acetyl CoA
yn pennu cyflymder yr holl rai dilynol
adweithiau synthesis lc
mae sitrad yn actifadu ensym mewn cytosol
carboxylase asetyl CoA. Malonyl CoA yn
yn ei dro yn atal trosglwyddo uwch
asidau brasterog o cytosol i fatrics
gweithgaredd atal mitochondria
CoA asetyl allanol: carnitine acyltransferase,
a thrwy hynny ddiffodd ocsidiad uwch
asidau brasterog.
Acaloyl-CoA Oxaloacetate →
Citrate HS-CoA

HSCOA ATP Citrate → Acetyl-CoA ADP Pi Oxaloacetate
Asetyl-CoA
yn y cytoplasm yn gwasanaethu fel y swbstrad cychwynnol
i mewn ar gyfer synthesis lcd, ac oxaloacetate yn
mae cytosol yn cael ei drawsnewid yn
y canlyniad y mae pyruvate yn cael ei ffurfio.
Biosynthesis colesterol
Mae biosynthesis colesterol yn digwydd yn y reticulum endoplasmig. Ffynhonnell yr holl atomau carbon yn y moleciwl yw asetyl-SCoA, sy'n dod yma o mitocondria fel rhan o sitrad, fel yn y synthesis o asidau brasterog. Mae'r biosynthesis colesterol yn defnyddio 18 moleciwl ATP a 13 moleciwl NADPH.
Mae ffurfio colesterol yn digwydd mewn mwy na 30 o ymatebion, y gellir eu grwpio mewn sawl cam.
1. Synthesis asid mevalonig.
Mae'r ddau adwaith synthesis cyntaf yn cyd-fynd â'r adweithiau ketogenesis, ond ar ôl synthesis 3-hydroxy-3-methylglutaryl-ScoA, mae'r ensym yn mynd i mewn hydroxymethyl-glutaryl-ScoA reductase (HMG-SCOA reductase), gan ffurfio asid mevalonig.
 |